
 (6 оценок, среднее: 4,67 из 5)
(6 оценок, среднее: 4,67 из 5)
Учебная работа № 1411. Математическое моделирование процесса триплеттриплетного переноса энергии
Безызлучательный перенос энергии триплетного возбуждения между молекулами – проблема весьма актуальная, поскольку этот процесс лежит в основе многих биологических процессов (фотосинтез), находит широкое применение в медицине (фотодинамическая терапия рака) и технике (лазеры на красителях). В связи с этим, изучение основных закономерностей межмолекулярного триплеттриплетного переноса энергии представляет определённый интерес для науки.
Основные параметры этого процесса установлены при исследовании фосфоресценции донора в отсутствие и в присутствие акцептора в твёрдых растворах. Для однокомпонентных растворов кинетика заселения и распада триплетных состояний хорошо изучена теоретически и экспериментально [ ]. Для двухкомпонентных растворов, которые используются для наблюдения межмолекулярного триплеттриплетного переноса энергии, теоретического исследования законов разгорания и затухания сенсибилизированной фосфоресценции в литературе не обнаружено.
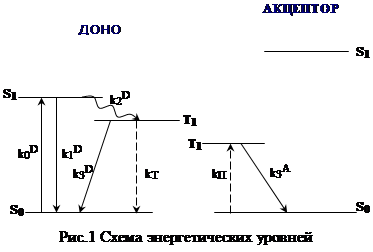 Для рассмотрения кинетики накопления триплетных молекул акцептора использовалась трехуровневая схема для молекул донора и двухуровневая для молекул акцептора (рис.1). Константы скоростей соответствующих переходов обозначены следующим образом (в нашем случае константа перехода есть сумма констант излучательного и безызлучательного переходов ):
Для рассмотрения кинетики накопления триплетных молекул акцептора использовалась трехуровневая схема для молекул донора и двухуровневая для молекул акцептора (рис.1). Константы скоростей соответствующих переходов обозначены следующим образом (в нашем случае константа перехода есть сумма констант излучательного и безызлучательного переходов ):
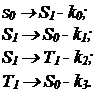 Концентрация молекул в состоянии S0 обозначена через n0, в состоянии S1 n1, в T1 n2.
Концентрация молекул в состоянии S0 обозначена через n0, в состоянии S1 n1, в T1 n2.
Кинетика накопления триплетных молекул донора в присутствии акцептора описана уравнениями (1)(3), кинетика накопления триплетных молекул акцептора – (4)(5).
![]() ; (1)
; (1)
![]() ; (2)
; (2)
![]() ; (3)
; (3)
![]() ; (4)
; (4)
![]() , (5)
, (5)
где индексы A и D указывают на то, что данная величина относится к молекулам акцептора или донора соответственно; N – общее число молекул в растворе, участвующих в данном процессе; kT константа тушения триплетного состояния донора за счёт переноса энергии на акцептор; kП – константа перехода молекул акцептора из основного состояния в триплетное в результате переноса энергии; k0=IВR (IВ интенсивность возбуждающего излучения; R – константа).
Кинетика дезактивации триплетного состояния донора в присутствии молекул акцептора описана уравнением (6), кинетика дезактивации триплетных молекул акцептора – (7):
![]() , (6)
, (6)
![]() . (7)
. (7)
Константа тушения триплетных молекул донора kT связана с константой перехода молекул акцептора kП следующим образом. Число триплетных молекул донора, перешедших за время dt в основное состояние за счет переноса энергии равно числу молекул акцептора, перешедших в триплетное:
![]() . (8)
. (8)
Решения уравнений (1)(3) и (4)(5) показали, что закон накопления триплетных молекул акцептора, как и закон накопления триплетных молекул донора носят экспоненциальный характер. Значения, к которым стремятся при насыщении концентрации триплетных молекул донора ![]() и акцептора
и акцептора ![]() (условия стационарного возбуждения) различные:
(условия стационарного возбуждения) различные:
 , (9)
, (9)
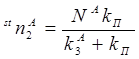 . (10)
. (10)
Существенным образом различаются также времена накопления числа триплетных молекул донора и акцептора.
Дезактивация триплетных состояний молекул донора и акцептора после прекращения возбуждения происходят по экспонентам, с соответствующими временами, отличными от времен накопления. Причем как для донора, так и для акцептора, время накопления всегда меньше или равно времени дезактивации.
В отсутствие реабсорбции излучения стационарная интенсивность фосфоресценции пропорциональна концентрации триплетных молекул,
Зависимость стационарной концентрации триплетных молекул акцептора энергии от мощности возбуждения
Зависимость интенсивности сенсибилизированной фосфоресценции (стационарной) от мощности возбуждения, как следует из (13) и (11), можно представить в виде
![]() , (16)
, (16)
где ![]() постоянные величины. В выражении (16) от мощности возбуждения зависит величина
постоянные величины. В выражении (16) от мощности возбуждения зависит величина![]() .
.
Решение системы уравнений (1а)(4а) дает зависимость ![]() от возбуждающего света
от возбуждающего света
![]() . (17)
. (17)
Введя соответствующие обозначения: ![]() ,
, ![]()
![]() и подставляя (17) в (16), окончательно получим
и подставляя (17) в (16), окончательно получим
![]() . (18)
. (18)
Таким образом, формально зависимость интенсивности СФ от мощности возбуждения (18) совпадает с зависимостью интенсивности обычной фосфоресценции от интенсивности возбуждения [2.3.4; 2.3.5].
Значение стационарного числа триплетных молекул акцептора получаем из (12) при t ®¥
![]() . (19)
. (19)
Учитывая, что
![]() и
и ![]() , (20)
, (20)
получаем
![]() . (21)
. (21)
Таким образом, подставляя (20) в (21), имеем
![]() . (22)
. (22)
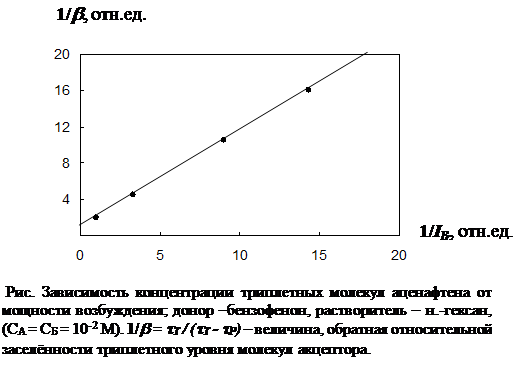
Для проверки полученных теоретических выводов было проведено экспериментальное исследование зависимости концентрации триплетных молекул аценафтена (акцептор энергии) от интенсивности возбуждающего света. Интенсивность возбуждающего света изменялась с помощью нейтральных фильтров (калиброванных металлических сеток), а концентрация триплетных молекул определялась по формуле (22). Экспериментальные результаты приведены на рис.2.3.2, где по оси абсцисс отложена величина, обратная интенсивности возбуждающего света IВ. За единицу приято максимальное возбуждение, которое соответствует относительной концентрации триплетных молекул 0.5. По оси ординат отложена величина, обратная относительной заселённости триплетного уровня молекул акцептора. Как видно из рисунка, экспериментальные точки хорошо укладываются на прямую, что согласуется с выражением (18).
 |
Полученное равенство (22) не только позволяет найти число триплетных молекул акцептора из кинетических параметров, но и определить константу перехода молекул акцептора в триплетное состояние
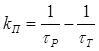 . (23)
. (23)
На основании всего вышесказанного можно сделать следующие выводы.
Для донорноакцептроных пар, удовлетворяющим условиям (8) –(10), закон разгорания сенсибилизированной фосфоресценции имеет экспоненциальный характер. Определенные из кинетических экспериментов параметры – время разгорания и затухания сенсибилизированной фосфоресценции – позволяют определить по формуле (22) долю молекул акцептора в триплетном состоянии от общего числа, участвующих в триплеттриплетном переносе энергии (b).
Формула (22) для определения числа триплетных молекул акцептора совпадает с формулой, полученной ранее М.В.Алфимовым с сотрудниками, для определения числа молекул в триплетном состоянии при возбуждении быстрыми электронами [2.3.1]. Однако время разгорания сенсибилизированной фосфоресценции зависит не только от интенсивности возбуждения, но и от константы переноса энергии в донорноакцепторной паре.
Зависимость интенсивности сенсибилизированной фосфоресценции от мощности возбуждения в указанном выше приближении определяется выражением (18), которое по форме совпадает с зависимостью интенсивности фосфоресценции от мощности возбуждения при фотовозбуждении.
Выражение (23) позволяет установить константу перехода молекул акцептора из основного состояния в триплетное в результате переноса энергии.